
Midgut NPF suppresses sugar intake and energy breakdown
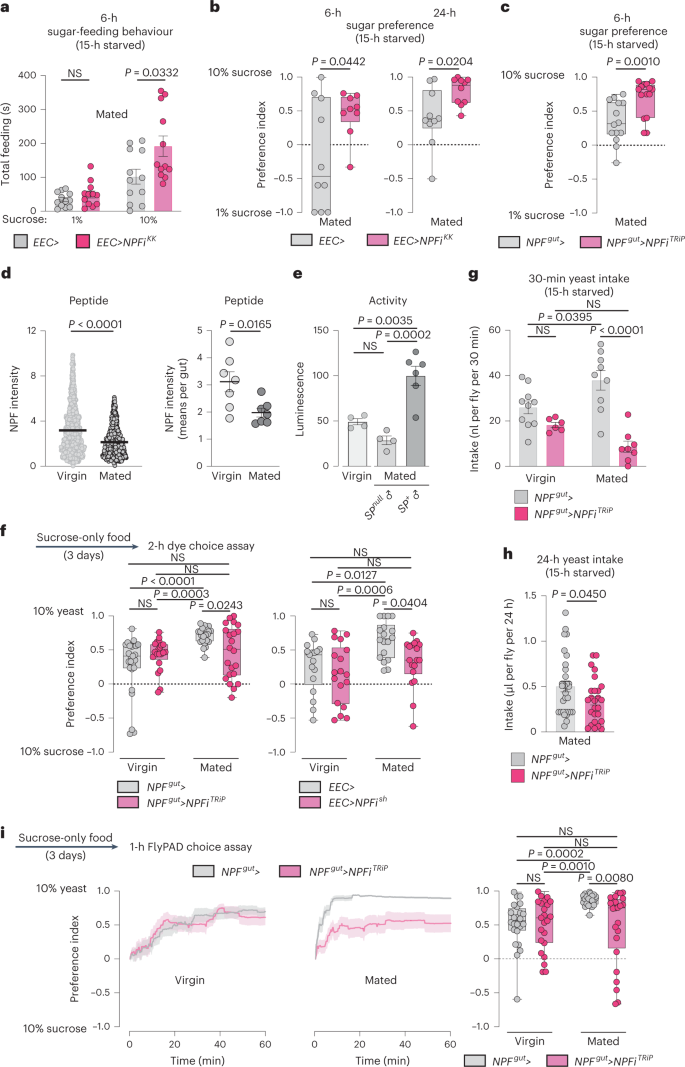
To identify gut-derived hormones and nutrient-sensing mechanisms that regulate feeding, we performed an in vivo RNA-interference screen of secreted factors and receptors in adult Drosophila. We focused on the EECs, which produce a variety of factors that play key roles in the coordination of food intake and metabolism12,18,20,21. We examined the effect of adult-restricted, EEC-specific gene knockdown on the sugar-water feeding behaviour of fed males and females (Fig. 1a) using the fly liquid-food interaction counter (FLIC) system, which allows automated monitoring of Drosophila feeding behaviours22. We used the driver voilà-GAL4 to target the RNAi effect to the EECs, in combination with ubiquitously expressed temperature-sensitive GAL80 (Tub-GAL80TS), together referred to as EEC> hereafter, which allowed us to induce gene silencing only in the adult stage18,20. Among our hits was the peptide NPF, knockdown of which increased the feeding time of mated females on sugar-only food while decreasing males’ sugar-interaction time (Fig. 1b and Extended Data Fig. 1a). To rule out contributions of the UAS transgene itself to the phenotype, we crossed the UAS-NPF-RNAiKK (NPFiKK) line to the control w1118 background. This genotype showed results similar to those seen with the driver control (Extended Data Fig. 1b). This suggests that lack of NPF production in the EECs of mated females enhances their interest in or motivation to feed on sugar.
a, Sugar feeding in mated females with RNAi-mediated knockdown of hormones and transporters in the EECs of the midgut. b, Total time feeding using FLIC; n = 16 EEC>, n = 9 EEC > NPFiKK. c,d, Consumption (c) of sugar+yeast food (9% sugar and 8% yeast) determined by dye assay, and of 10% sugar (d) measured by CAFÉ assay; c, n = 8 EEC> and EEC>NPFish; d, n = 8 EEC>, n = 9 EEC>NPFiKK. e, Conditional NPF knockdown with EEC> affects the EECs but not the CNS (brain and VNC); n = 6 biological replicates from tissues pooled from six animals for each condition. f,g, NPF immunostaining of CNS and midgut, quantified in g; n = 7 CNS, n = 6 midguts. Scale bars, 50 μm. h, Quantification of images represented in Extended Data Fig. 1i, immunostaining of NPF knockdown using the NPF> driver with pan-neuronal GAL80 (R57C10-GAL80; NPF>– together, NPFgut>) of midgut EECs and CNS; n = 7 tissues each. i, Intake measured by dye-consumption and CAFÉ assays. Left n = 10 NPFgut>, n = 9 NPFgut>NPFiTRiP; right n = 14 NPFgut>, n = 15 NPFgut>NPFiTRiP. j,k, Consumption and glycaemic levels after injection of NPF peptide into the haemolymph. j, n = 9 each. k, n = 11 each. l, Thirty-minute food intake measured by dye assay during activation of NPF+ EECs using the heat-sensitive TrpA1 channel; n = 10 NPFgut>, n = 8 NPFgut>TrpA1, n = 9 NPFgut>TrpA1, NPFiTRiP. m, Survival under starvation. n, TAG levels; n = 8 fed EEC>, n = 10 fed EEC>NPFiKK, n = 9 starved EEC>, n = 9 starved EEC>NPFiKK. All animals were mated females. Bars represent mean ± s.e.m. NS, not significant. b–d,i, Two-tailed unpaired Student’s t-test. e,g,h,n, Two-tailed unpaired Mann–Whitney U-test. j–l, One-way ANOVA with Tukey’s multiple-comparisons test. m, Kaplan–Meier log-rank tests.
Source data
Knockdown of gut NPF throughout development has recently been associated with increased consumption of food containing both sugar and yeast in virgin female adults21. We therefore analyzed whether EEC-derived NPF also regulates intake of sugar+yeast food in mated females. We measured short-term (30-minute) food intake using a dye-consumption assay with standard adult fly food23 containing both sugar (9%) and yeast (8%). To measure short-term intake, we preconditioned animals by fasting them for 15 hours to increase consumption. We confirmed that, like virgins, mated females with adult-restricted EEC knockdown of NPF also consumed significantly more sugar+yeast food than controls (Fig. 1c and Extended Data Fig. 1c). We also applied the capillary feeder (CAFÉ) assay24 to quantify sugar intake over a longer period. Before this assay as well, we exposed animals to a 15-hour period of fasting to enhance consumption. Mated females with adult-restricted EEC knockdown of NPF consumed significantly more sugar over the 6-hour period than controls (Fig. 1d and Extended Data Fig. 1d). Since the sugar-feeding phenotype was observed in animals whether they were fully fed or preconditioned by 15-hour fasting (Fig. 1b,c,d), we chose to use animals that were 15-hour fasted for consistency in the following feeding assays, since this allows robust measurements in short-term food intake assays as well as longer-term feeding assays. Conversely, in males, loss of NPF in the EECs led to reduced food consumption over an even longer period of 24 hours (Extended Data Fig. 1e). Together, these results indicate that gut-derived NPF suppresses sugar intake in females.
To attribute these effects specifically to EEC-derived NPF, we measured the expression of NPF in dissected midguts and central nervous systems (CNS; brain and ventral nerve cord, VNC). NPF transcript levels were strongly reduced in the female midgut when EEC> was used to drive knockdown of NPF, whereas expression in the CNS was unaltered, which was confirmed by immunostainings (Fig. 1e,f,g and Extended Data Fig. 1f,g). To further support this, we used a second driver, NPF::2A::GAL4 (NPF>), a CRISPR-mediated insertion of T2A::GAL4 into the native NPF locus that drives GAL4 expression in only NPF-producing cells25. Since NPF is expressed in both neurons and EECs, we used pan-neuronal R57C10-GAL80, an optimized nSyb-GAL80 variant that suppresses neuronal GAL4 (ref. 18), to suppress the GAL4 activity of NPF> in the nervous system. We confirmed that this driver combination (R57C10-GAL80; NPF>), referred to hereafter as NPFgut>, efficiently knocks NPF down in the midgut without affecting CNS expression (Fig. 1h and Extended Data Fig. 1h,i). Knockdown of NPF using this gut NPF-specific driver caused a marked increase in intake of sugar+yeast food measured over 30 minutes and in consumption of sugar-only medium measured over 6 hours after 15-hour starvation (Fig. 1i and Extended Data Fig. 1j). Taken together, these data indicate that EEC-specific loss of NPF is responsible for the observed feeding phenotypes and indicate that gut NPF acts as a satiety signal that inhibits sugar consumption.
To examine the ability of NPF to promote satiety, we injected synthetic NPF peptide into circulation in mated females. EEC-specific knockdown of NPF induced hyperphagia, which was blocked by NPF injection (Fig. 1j and Extended Data Fig. 1k). NPF injection did not affect haemolymph sugar levels (Fig. 1k), indicating that the observed feeding effect was not a consequence of alterations in glycaemic levels. Next, we expressed the thermosensitive Transient receptor potential A1 (TrpA1) cation channel26 in the NPF+ EECs to enable induction of NPF release. Incubation at 29 °C, which induces TrpA1-mediated peptide release, inhibited food intake, an effect that was abolished by simultaneous NPF knockdown (Fig. 1l). These NPF-induced changes in food intake were likewise not associated with altered triacylglyceride (TAG) or circulating sugar levels (Extended Data Fig. 1l,m), supporting a direct role for gut-derived NPF in governing feeding behaviour, rather than effects of NPF on metabolism that then lead secondarily to altered behaviour. Together, these results indicate that NPF from these EECs is both necessary and sufficient to inhibit food intake and prevent food overconsumption.
Feeding behaviours are tightly coordinated with physiology to maintain metabolic balance. Our findings indicate that NPF acts as a satiety signal, which suggests that it should act after a meal. In this scenario NPF would be expected to promote storage and inhibit mobilization of energy. As an indirect measure of energy storage and mobilization, we first assessed animals’ starvation resistance. NPF knockdown in the EECs throughout development (voilà> without Tub-GAL80TS (Fig. 1m)), as well as adult-restricted RNAi (Extended Data Fig. 2a,b,c), led to a decrease in starvation resistance in females but not males, in line with a recent study linking gut NPF to metabolic programs associated with energy storage21. Consistent with their shortened starvation survival, we found that females with constitutive (Extended Data Fig. 2d) or adult-restricted (Fig. 1n and Extended Data Fig. 2e) EEC knockdown of NPF showed a decrease in both TAG and glycogen levels, whereas TAG levels were not affected by EEC-specific NPF loss in males. These observations suggest that although NPF does affect metabolism in the adult stage, it also regulates early life history in ways that affect the adult. We found that haemolymph glucose levels increased more after re-feeding in animals with EEC suppression of NPF, consistent with their increased sugar consumption (Extended Data Fig. 2f), and showing that the mechanisms of sugar absorption and transport into circulation are functional. Together our findings indicate that, in addition to the metabolic findings described recently21, a main function of EEC-derived NPF in the adult stage is the regulation of feeding, particularly the inhibition of sugar intake in mated females.
Gut NPF suppresses sugar intake and regulates food choice
Our findings suggest that NPF acts as a sugar-satiety signal. To test this hypothesis directly, we examined whether gut NPF affected animals’ preference for dietary sugar when they were given the choice between two different sucrose concentrations (1 and 10%). NPF knockdown in the EECs increased feeding and preference for 10% sugar in mated females (Fig. 2a,b,c and Extended Data Fig. 3a,b). These results indicate that NPF is part of a postingestion sugar-sensing mechanism required to decrease sugar appetite.
a, Time spent feeding on 1 or 10% sucrose using FLIC; all n = 12 animals. b,c, Preference between 1 versus 10% sugar measured over 6 or 24 hours by CAFÉ assay. b, 6-hour preference: n = 10 EEC>, n = 9 EEC>NPFiKK. 24-hour preference: n = 10 each. c, n = 14 NPFgut>, n = 15 NPFgut>NPFiTRiP. d, Midgut NPF staining intensity in fed females on a per-cell basis and on a per-gut basis; n = 703 cells from seven guts for virgins, n = 883 cells from seven guts from mated females. e, NPF-cell activity measured in dissected midguts (two midguts per replicate) using a luciferase-based CaLexA calcium-reporter system (NPF>LexA::NFAT::VP16; LexAop-luciferase); n = 4 from fed virgins, n = 4 from fed females mated to SP-deficient males (SP0/Df(3L)delta130), n = 6 from fed females mated to SP+ males. f, Consumption preference for 10% sucrose versus 10% yeast after 3 days of yeast deprivation (3 days on sucrose-only medium) using two-choice dye assay; n = 25 NPFgut> virgins, n = 23 NPFgut>NPFiTRiP virgins, n = 24 NPFgut> mated females, n = 22 NPFgut>NPFiTRiP mated females, n = 17 EEC> virgins, n = 18 EEC>NPFish virgins, n = 18 EEC> mated females, n = 18 EEC>NPFish mated females. g, Yeast consumption determined by dye assay; virgins n = 10 NPFgut> virgins, n = 6 NPFgut>NPFiTRiP virgins, n = 9 NPFgut> mated females, n = 8 NPFgut>NPFiTRiP mated females. h, Yeast intake determined by CAFÉ; n = 32 NPFgut>, n = 26 NPFgut>NPFiTRiP. i, Cumulative behavioural preference of females for sucrose versus yeast monitored using flyPAD. Lines represent means, and shading indicates s.e.m.; n = 24 NPFgut> virgins, n = 23 NPFgut>NPFiTRiP virgins, n = 24 NPFgut> mated females, n = 24 NPFgut>NPFiTRiP mated females. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not significant. a,c,d (left), Two-tailed unpaired Mann–Whitney U-test. b,d (right), h, Two-tailed unpaired Student’s t-test. e,f (right), g, One-way ANOVA with Tukey’s post hoc test. f (left), i, Kruskal–Wallis nonparametric ANOVA with Dunn’s multiple-comparisons test.
Source data
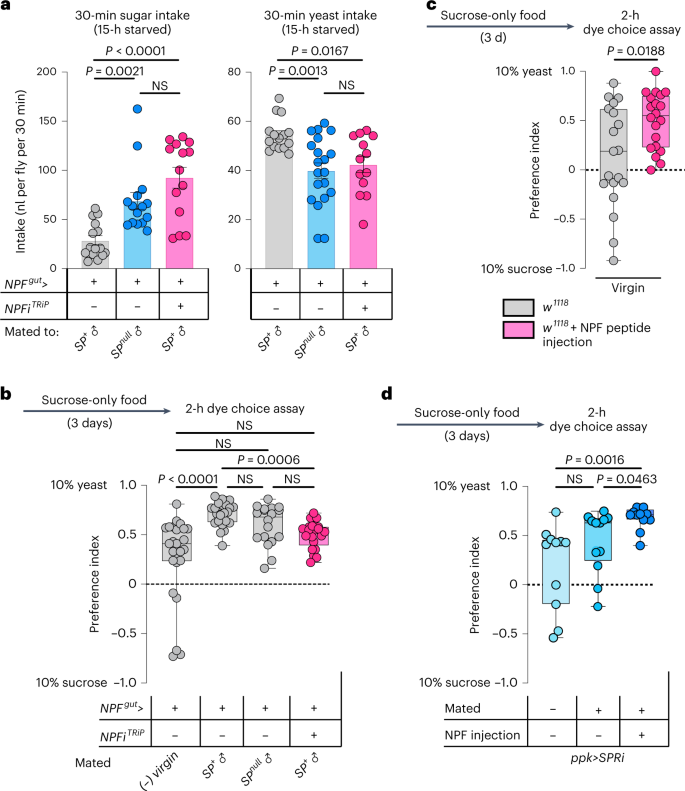
Given that females increase their preference for protein-rich food after mating to meet the metabolic requirements of egg production6, we speculated that gut NPF might be important to reduce sugar appetite and increase intake of protein-rich food in mated females. Postmating sex peptide (SP) signalling within the female induces an increased preference for yeast6, and this peptide has also been shown to potentiate NPF release from the midgut17. We confirmed that mating induces NPF secretion from the midgut by measuring EEC NPF protein levels. After mating, NPF peptide levels were reduced in the midgut, consistent with increased release (Fig. 2d). Using a luciferase-based CaLexA reporter27, in which calcium induces the expression of luciferase, we found that NPF+ EECs showed increased calcium-reporter activity after mating in an SP-dependent manner (Fig. 2e). We therefore proposed that gut-derived NPF might be involved in mediating the SP-induced increase in protein consumption in mated females. To test this possibility, we investigated whether NPF affects yeast preference by using a two-choice dye-based consumption assay to measure preference between sugar or protein-rich yeast food6. Animals were deprived of protein for 3 days before the experiment by keeping them on sucrose-only food to increase their preference for yeast food6, making any reduction in this preference easier to observe. We observed that mating increased control females’ preference for yeast food after this treatment (Fig. 2f), consistent with previous findings6. However, animals with EEC-specific NPF loss displayed a reduced preference for yeast food that did not significantly increase after mating. This indicates that EEC-derived NPF is required to inhibit sugar intake in mated females, thereby promoting consumption of protein-rich food. Consistent with this, control animals increased their yeast consumption after mating, whereas mating did not significantly increase yeast consumption in females lacking gut NPF that consumed less yeast (Fig. 2g,h). To further test our conjecture that gut NPF is involved in mating-induced yeast consumption, we used a second automated behaviour-monitoring apparatus, the flyPAD28, to measure feeding preference in a two-choice assay. Behavioural results obtained with this assay indicate that gut NPF is important for the mating-induced increase in preference for yeast (Fig. 2i). Together, these data indicate that NPF from the midgut is involved in promoting yeast intake in mated females, an effect triggered by SP signalling6. Consistent with this notion, females with EEC-specific NPF knockdown mated to SP+ males displayed a consumption pattern similar to that of control females mated to SP-mutant males: both consumed more sugar and less protein than control females mated to SP+ males (Fig. 3a). Control females mated to SP-mutant males showed a yeast-preference phenotype that was intermediate between those of virgin females and females mated to SP+ males, as previously reported6. Females with EEC-specific NPF knockdown mated to SP+ males displayed lower yeast preference than control females mated to SP+ males, and their yeast preference was not significantly different from that of virgin females (Fig. 3b). Thus, females upregulate their protein intake after mating in a partially SP-dependent manner, and our results indicate that NPF from the EECs is involved in mediating this SP-induced shift in food choice, independently of juvenile hormone (Extended Data Fig. 3c), which is known to affect gut remodelling after mating29. We therefore rationalized that injection of NPF into virgin females should induce an increase in their yeast preference. As expected, virgin females injected with NPF peptide exhibited an increased preference for dietary yeast (Fig. 3c).
a, Intake of sucrose or yeast measured by dye assay in females mated to SP-producing or SP-deficient males (SP0/Df(3L)delta130). Sucrose, n = 15 NPFgut> females mated to SP+ males, n = 15 NPFgut> females mated to SP-mutant (SPnull) males, n = 14 NPFgut>NPFiTRiP females mated to SP+ males. Yeast, n = 15 NPFgut> females mated to SP+ males, n = 20 NPFgut> females mated to SP-mutant males, n = 13 NPFgut>NPFiTRiP females mated to SP+ males. b, Consumption preference using two-choice dye assay; n = 25 NPFgut> virgins, n = 24 NPFgut> females mated to SP+ males, n = 18 NPFgut> females mated to SP-null males, n = 21 NPFgut>NPFiTRiP females mated to SP+ males. c,d, Consumption preference of w1118 virgin females with or without NPF injection (c) and virgin and mated females (d) (with and without NPF injections) with knockdown of SPR in Ppk+ neurons (ppk>SPRi) using two-choice dye assay. c, n = 19 w1118 virgins, n = 20 w1118 virgins with NPF injection. d, n = 11 virgin ppk>SPRi females, n = 12 mated ppk>SPRi females, n = 12 mated ppk>SPRi females with NPF injection. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not significant. a (left), b,d, One-way Kruskal–Wallis ANOVA with Dunn’s multiple-comparisons test. a, Right, one-way ANOVA with Dunnett’s multiple-comparisons test. c, Two-tailed unpaired Mann–Whitney U-test.
Source data
The induction of yeast preference and the stimulation of gut NPF release after mating are both triggered by SP receptor (SPR) activity in reproductive-tract Ppk+ neurons6,17. We found that mating did not significantly upregulate yeast preference in females with SPR knockdown in the Ppk+ neurons (Fig. 3d), confirming that SP/SPR signalling is important for the preference change. However, injection of NPF was still able to increase the yeast preference of these mated females (Fig. 3d), suggesting that gut NPF acts downstream of SP-SPR signalling in mated females to regulate food choice.
Sut2 in NPF+ EECs regulates NPF release and food choice
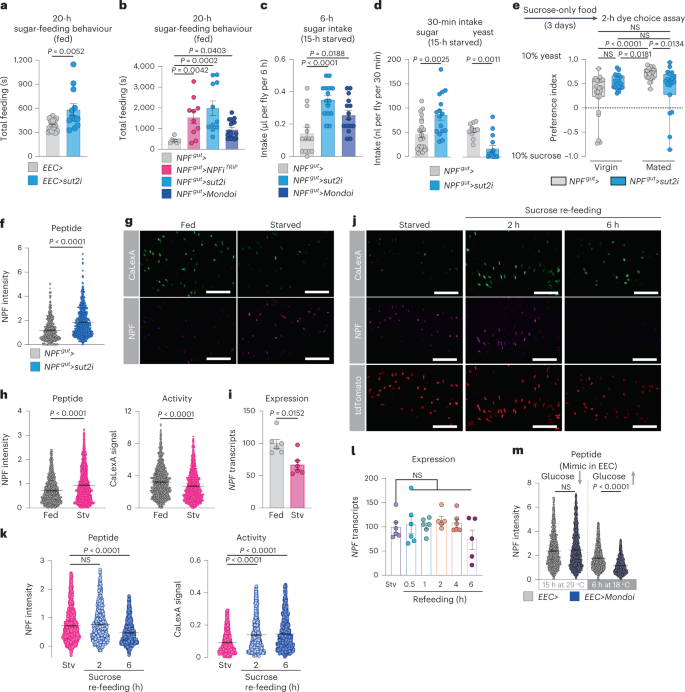
Given these results indicating that NPF acts as a mediator of sugar satiety, we asked whether NPF-producing EECs might be activated by sucrose ingestion. In our initial analysis of a collection of RNAi lines (Fig. 1a), we found that knockdown of sugar transporter 2 (sut2), a member of the glucose-transporter class of solute carrier (SLC) proteins, in the EECs of fed mated females increased their sugar-feeding behaviour (Fig. 4a). Loss of sut2 in NPF+ EECs increased animals’ sugar-feeding behaviour and sugar intake, similar to the effects observed in animals with knockdown of NPF itself (Fig. 4b,c and Extended Data Fig. 3d), suggesting that Sut2 might be required as part of a mechanism governing NPF production or release.
a,b, Feeding time measured using FLIC. a, n = 16 EEC>, n = 12 EEC>sut2i. b, n = 7 NPFgut>, n = 10 NPFgut>NPFiTRiP, n = 11 NPFgut>sut2i, n = 17 NPFgut>Mondoi. c, Sugar consumption measured by CAFÉ assay; n = 14 NPFgut>, n = 15 NPFgut>sut2i, n = 15 NPFgut>Mondoi. d, Consumption of sucrose or yeast measured by dye assay. Sucrose n = 22 NPFgut>, n = 16 NPFgut>sut2i. Yeast n = 10 NPFgut>, n = 15 NPFgut>sut2i. e, Consumption preference measured by two-choice dye-consumption assay. Virgins, n = 25 NPFgut>, n = 17 NPFgut>sut2i; mated n = 24 NPFgut>, n = 19 NPFgut>sut2i. f, Midgut NPF staining with sut2 knockdown; n = 571 cells from six guts for NPFgut>, n = 625 cells (five guts) for NPFgut>sut2i. g,h, Representative images of midgut NPF-cell activity (g), quantified in h; left shows n = 1,167 cells from eight fed guts, n = 1,394 cells from nine starved (stv) guts; right shows n = 1,209 cells from eight fed guts, n = 1,405 cells from nine starved guts. i, Midgut NPF expression; n = 6 for stv, 0.5 h, 1 h, 4 h and n = 5 for 2 h, 6 h, each samples of five guts. j, Representative images of NPF staining and cell activity in 24-h starved, 2-h sugar re-fed and 6-h sugar re-fed mated females. All measured cells are marked with tdTomato. k, Quantification of j. Eight guts per condition. Left, n = 1,297, 1,038 and 1,216 cells from starved, 2-h and 6-h re-fed animals. Right, n = 1,141, 954 and 1,151 cells from starved, 2-h re-fed and 6-h re-fed animals. l, Midgut NPF expression in mated females after re-feeding following 24-hour starvation (Stv, no re-feeding); n = 6 replicates of six midguts except n = 5 replicates for 2 and 6 h. m, NPF staining in mated females’ midguts after 15 hours’ knockdown of Mondo (29 °C inactivation of GAL80TS) and following subsequent 6-h derepression of Mondo (18 °C). 15 h at 29 °C: n = 1,595 cells/12 guts for EEC>, n = 1,840 cells/14 guts for EEC>Mondoi; 6 h at 18 °C: n = 1,948 cells/17 guts for EEC>, n = 2,194 cells/18 guts for EEC>Mondoi. All animals were mated females, except in e. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not significant. a,d (left), Two-tailed unpaired Student’s t-test. b,e,k, One-way Kruskal–Wallis ANOVA with Dunn’s multiple-comparisons test. c,l, One-way ANOVA with Dunnett’s multiple-comparisons test. d (right), f,h,i,m, Two-tailed unpaired Mann–Whitney U-test. Scale bars, 50 μm.
Source data
In Drosophila and mammals, the sugar-responsive transcription factor Mondo/ChREBP (carbohydrate-responsive-element-binding protein) contributes to many of the cellular responses to sugar30. To probe the molecular sugar-sensing mechanisms regulating NPF, we silenced Mondo specifically in NPF+ EECs of mated females and found that this manipulation increased both sugar-feeding behaviour and sugar intake, although not as dramatically as the loss of NPF or sut2 (Fig. 4b,c). This suggests that although other mechanisms are probably involved, Mondo/ChREBP-mediated sugar sensing may contribute to NPF regulation in EECs.
Because NPF loss led to increased sugar intake and decreased protein feeding, we investigated whether Sut2 also affects sugar versus protein intake in mated females. We found that knockdown of sut2 in NPF+ EECs led to a strongly increased intake of sucrose and a marked decrease in consumption of yeast when animals were presented with these foods separately (Fig. 4d). Similarly, when given a choice between these two foods, mated females with knockdown of sut2 in NPF+ EECs displayed a reduced preference for dietary yeast (Fig. 4e). Although Sut1 has been linked to NPF secretion in virgins21, we did not observe significant changes in yeast preference in virgins or mated females with sut1 knockdown in NPF+ EECs (Extended Data Fig. 3e). These results indicate that loss of sut2 in NPF+ EECs shifts consumption towards sugar-rich food, similar to knockdown of NPF in these same gut cells, consistent with a role for Sut2 in regulating NPF production or release. Knockdown of sut2 in NPF+ EECs led to a strong intracellular accumulation of NPF peptide, even though NPF transcript levels were reduced (Fig. 4f and Extended Data Fig. 3f,g), suggesting that sut2 loss in NPF+ EECs leads to NPF retention and thus that Sut2 is required for normal NPF expression and release. We also found that sut2 transcript levels in the entire midgut were strongly reduced by knockdown targeted only at the NPF+ gut endocrine cells (Extended Data Fig. 3g), demonstrating that sut2 is predominantly expressed in NPF+ EECs.
To assess more directly whether sugar regulates NPF+ EECs, we exposed mated females to different nutritional conditions and observed their calcium-signalling history, using the CaLexA reporter system, in which green fluorescent protein (GFP) expression reflects calcium signalling31. After 24 hours of starvation, we observed decreased calcium-induced GFP signal and increased NPF peptide staining in the NPF+ EECs when measured on a per-cell basis, even though NPF transcript levels were reduced (Fig. 4g–i), indicating NPF retention. Although these measures were higher in starved animals on a per-cell basis (Fig. 4g,h), they were not significantly altered when analyzed on a one-mean-per-gut basis (Extended Data Fig. 3h). This might possibly reflect the inhibition of only a subpopulation of the NPF+ EECs by starvation, which could be masked by averaging all the cells, or that starvation longer than 24 hours is required for strong inhibition, as also found by a recent report showing that NPF release is reduced after 48 hours’ starvation21. Re-feeding with sucrose after starvation elicited a strong increase in calcium signalling within 2 hours in the NPF+ EECs, associated with a decrease in NPF peptide staining within 6 hours of re-feeding (Fig. 4j,k and Extended Data Fig. 3i), as also reported by an independent study21. Since midgut NPF transcript levels were unaltered under these conditions (Fig. 4l), these results indicate that midgut NPF+ cells are activated by dietary sugar, leading to their secretion of NPF peptide.
We then used genetic methods to mimic sugar sensing occurring in the EECs following a meal. We first induced RNAi-mediated silencing of Mondo/ChREBP by switching flies to 29 °C to inactivate GAL80TS for 15 hours, to reduce sugar sensing in the EECs. We then reactivated Mondo/ChREBP signalling, to mimic sugar-sensing occurring after a meal, by switching animals from 29 back to 18 °C to renature GAL80TS and thereby inactivate the RNAi effect. Reactivation of Mondo/ChREBP signalling in the EECs caused a decrease in NPF peptide levels in these cells without altering NPF expression (Fig. 4m and Extended Data Fig. 3j,k), indicating increased NPF secretion, consistent with the notion that sugar sensing in the EECs is associated with NPF release. Taken together, our findings indicate that, in mated females, sugar intake leads to EEC NPF release through a process requiring glucose-transporter-family protein Sut2 and involving Mondo/ChREBP-mediated sugar sensing.
NPF suppresses energy mobilization in adipose tissues
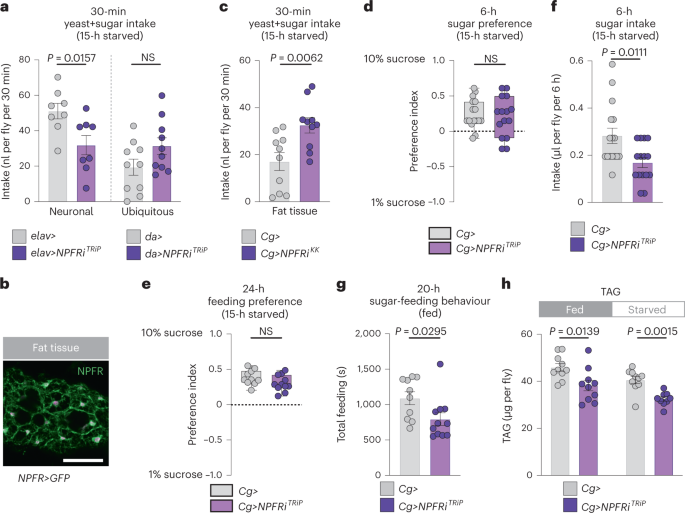
We then asked which target tissues might be involved in NPF-mediated appetite regulation by knocking down the NPF receptor (NPFR). Whereas EEC-specific NPF knockdown induced overfeeding (Fig. 1d), adult females with neuronal knockdown of NPFR driven by elav-GAL4 (elav>) showed decreased food intake (Fig. 5a), consistent with previous reports that neuronal NPF/NPFR signalling promotes feeding32 and indicating that other tissues mediate the downregulation of sugar intake induced by gut NPF. Global knockdown of NPFR driven by daughterless-GAL4 (da>) led to a feeding phenotype intermediate between those observed with gut or neuronal NPF signalling loss (Fig. 5a), probably reflecting opposing effects on feeding of NPF signalling within different organs.
a, Food intake measured by dye in animals with knockdown of NPFR in the nervous system (elav>) or the entire body (da>) measured by dye assay; n = 8 elav>, n = 8 elav>NPFRiTRiP, n = 10 da>, n = 10 da>NPFRiTRiP. b, Fat-body immunostaining of NPFR>mCD8::GFP reporter. Scale bar, 50 μm c, Food intake determined by dye assay in fat-body NPFR-knockdown animals; both n = 10. d, Consumption preference for 1 versus 10% sucrose, measured by CAFÉ assay; n = 16 Cg>, n = 15 Cg>NPFRiTRiP. e, Behavioural preference for interacting with 1 versus 10% dietary sucrose measured by FLIC; n = 10 Cg>, n = 11 Cg>NPFRiTRiP. f, Sucrose intake by CAFÉ assay; n = 16 Cg>, n = 15 Cg>NPFRiTRiP. g, Time spent feeding on sucrose using FLIC; n = 10 Cg>, n = 11 Cg>NPFRiTRiP. h, Whole-body TAG levels in fed and 15-hour-starved fat-body NPFR-knockdown animals. All n = 10 except n = 9 starved Cg>NPFRiTRiP. All animals were mated females. Bars represent mean ± s.em. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not significant. a,c–e,h (left), Two-tailed unpaired Student’s t-test. f–h (right), Two-tailed unpaired Mann–Whitney U-test.
Source data
To assess receptor expression in other target tissues, we used a CRISPR-mediated knock-in of T2A::GAL4 into the native NPFR locus (NPFR::T2A::GAL4, hereafter NPFR>)33 to express UAS-mCD8::GFP. We observed reporter expression in the fat body (Fig. 5b), a tissue analogous to adipose tissue and liver in mammals. Although fat-body-specific NPFR knockdown driven by Cg-GAL4 (Cg>) in adult females did lead to increased short-term intake of food containing both sugar and yeast, it did not increase preference for sugar (Fig. 5c–e and Extended Data Fig. 4a). Indeed, suppression of NPFR in the fat body led to decreased sucrose intake and sugar-feeding behaviour (Fig. 5f,g). Thus, fat-body NPFR signalling does not appear to underlie the specific feeding phenotypes observed with gut NPF loss. We next asked whether fat-body NPFR mediates the effects of gut NPF on metabolism. Like animals with gut-specific NPF knockdown, fat-body NPFR knockdown animals were more sensitive to starvation and displayed metabolic phenotypes similar to those seen with loss of gut NPF (Fig. 5h and Extended Data Fig. 4b,c). These findings indicate that gut-derived NPF acts on NPFR in the fat body as part of a metabolic pathway that maintains energy homeostasis.
NPF regulates food choice through glucagon-like signalling
Our experiments indicate that gut NPF signalling regulates sugar appetite via tissues other than the CNS and fat body. In Drosophila, the brain cells that produce insulin express NPFR21 and these cells also regulate aspects of feeding and satiety34. However, knockdown of NPFR in the insulin-producing cells (IPCs) did not change preference for yeast versus sugar in mated females (Extended Data Fig. 5a), suggesting that gut-derived NPF does not act through insulin to modulate preference for dietary sugar and protein.
To identify the tissue mediating this effect, we examined NPFR expression in other tissues, which revealed expression of the receptor in the cells producing the glucagon-like factor AKH (Fig. 6a). AKH is released from the AKH-producing cells (APCs) during starvation and acts through its receptor, AkhR, on the fat body to promote the mobilization of stored energy, and it is also thought to act as a hunger signal to drive feeding behaviours18,31,35,36,37. However, whether AKH regulates sugar- or protein-specific feeding is unknown. We proposed that gut-derived NPF, released in response to sugar feeding, might suppress AKH release from the APCs in the fed state. Consistent with a recent report21, we found that knocking down NPFR in the APCs using AKH-GAL4 (AKH>) resulted in decreased AKH peptide levels within these cells in fed mated females (Fig. 6b), indicating that NPFR is required to suppress AKH release when the animal has ingested food. AKH promotes lipid and glycogen breakdown, and we therefore tested whether NPFR activity in the APCs regulates metabolism. As with knockdown of NPF in the midgut, adult females with knockdown of NPFR in the APCs showed reduced TAG and glycogen levels and increased susceptibility to starvation, as also reported recently21 and consistent with an increase in AKH signalling (Fig. 6c,d).
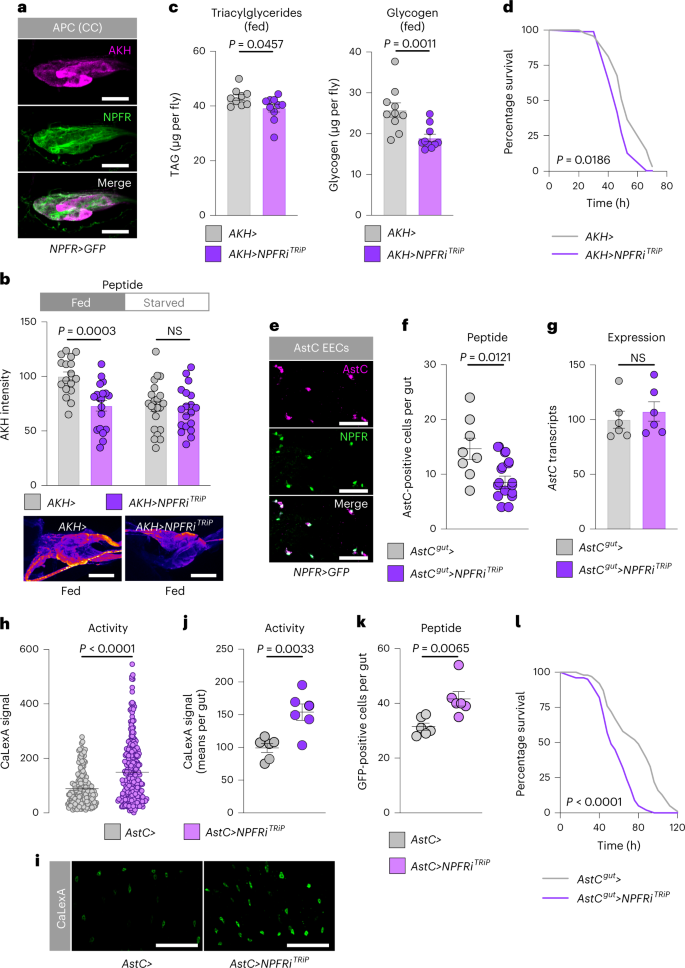
a, Immunohistochemistry of APCs shows NPFR>GFP reporter expression in APCs of mated females. Scale bars 20 μm. b, Quantification of AKH levels within the APCs of fed and 15-hour-starved mated females with and without NPFR knockdown in these cells; n = 17 fed AKH>, n = 19 fed AKH>NPFRiTRiP, n = 22 starved AKH>, n = 19 starved AKH>NPFRiTRiP. Representative images are shown below. Scale bars 20 μm. c, Metabolite levels in fed mated females. TAG n = 9 AKH>, n = 10 AKH>NPFRiTRiP; glycogen n = 10 AKH>, n = 10 AKH>NPFRiTRiP. d, Survival during starvation of mated females; n = 96 AKH>, n = 96 AKH>NPFRiTRiP. e, Immunohistochemistry of guts from mated female NPFR>GFP flies showing expression of NPFR reporter in AstC+ cells. Scale bars, 25 μm. f, Quantification of the number of midgut cells per gut that showed detectable AstC staining with and without NPFR knockdown in AstC+ EECs of fed mated females; n = 8 AstCgut> guts, n = 11 AstCgut>NPFRiTRiP. g, Midgut AstC transcript levels in fed mated females; each n = 6. h–k, AstC+ EEC-cell activity levels (h) with representative images (i) quantified on a per-gut basis (j) and the number of AstC+ EEC cells showing detectable GFP (k), measured by calcium-reporter system (AstC>LexA::NFAT::VP16; LexAop-GFP, denoted by AstC> in the figure) with or without NPFR knockdown in the AstC+ EECs in the midgut. h, n = 182 AstC> cells, n = 255 AstC>NPFR cells; j, each n = 6 guts; k, each n = 6 guts. Scale bars, 50 μm. l, Survival during starvation of mated females; n = 153 AstCgut> animals, n = 198 AstCgut>NPFRiTRiP. All animals were mated females. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not ignificant. b,c (left), f,g,j, Two-tailed unpaired Student’s t-test. d,l, Gehan–Breslow–Wilcoxon test. c (right), h,k, Two-tailed unpaired Mann–Whitney U-test.
Source data
We recently reported that the peptide hormone AstC released by midgut EECs promotes AKH release during starvation conditions18. We wondered whether NPF signalling might also inhibit AstC release from the midgut to suppress the activation of the AKH axis at multiple hierarchical levels. We found expression of NPFR reporter in AstC+ EECs (Fig. 6e), consistent with single-cell RNA-sequencing data38. We silenced NPFR expression specifically in AstC+ EECs using AstC-GAL4 (AstC>) with pan-neuronal GAL80 (R57C10-GAL80, AstC>–AstCgut> hereafter) to suppress nervous system GAL4 activity. This did not alter AstC expression, but it did lead to a reduction in the number of cells containing detectable AstC peptide (Fig. 6f,g), suggesting that EEC loss of NPFR cell-autonomously increases AstC release. Knockdown of NPFR with AstCgut> led to an increased number of active midgut AstC cells and to higher overall calcium-reporter activity (Fig. 6h–k), indicating that NPFR knockdown promotes AstC+ EEC activation. This indicates that the decreased number of AstC immune-positive cells in adult females with NPFR knockdown in the AstC+ EECs (Fig. 6f) is due to increased release of AstC peptide, which would be expected to promote AKH release, leading to more rapid depletion of energy stores and therefore reduced capacity to survive starvation18. Consistent with this, NPFR knockdown targeted to the AstC+ EECs led to a clear reduction in the capacity of these animals to survive starvation (Fig. 6l). Taken together, our results indicate that in response to sugar intake, gut-derived NPF inhibits the AKH axis at three levels: blocking the release of adipokineticotropic AstC from midgut EECs, blocking the release of AKH itself from the APCs and counteracting AKH’s effects on the fat body.
NPFR knockdown in the APCs was recently linked to increased consumption of food in virgin females21, an effect we confirmed in mated females (Fig. 7a). Next, we tested whether NPF regulates sugar- versus protein-specific feeding through NPFR in the APCs. Mated females with APC-specific NPFR knockdown exhibited elevated sugar-directed feeding behaviour, sugar consumption and preference for dietary sugar (Fig. 7b–e and Extended Data Fig. 5b–e), similar to animals with loss of NPF in the midgut. Next, we examined whether NPFR in the APCs might be involved in promoting yeast preference in mated females. Indeed, whereas APC knockdown of NPFR in virgin females did not detectably alter feeding preference, this manipulation had a strong effect in mated females (Fig. 7f and Extended Data Fig. 5f), similar to animals with knockdown of NPF in the gut, indicating that NPFR in the APCs is an important element for promoting consumption of protein-rich food in mated females. To determine whether AKH mediates the effects of NPFR loss on feeding, we examined the ability of AKH knockdown to rescue NPFR-RNAi’s sugar-overeating phenotype. We found that knockdown of AKH completely abolished this effect (Fig. 7g), suggesting that AKH is the primary factor mediating the feeding effects of NPFR signalling. Consistent with NPFR’s suppression of AKH release, AKH loss and NPFR knockdown induced opposite effects on sugar intake and yeast consumption (Fig. 7h,i). Together, our results indicate that, in mated females, gut-derived NPF acts on the APCs via NPFR to inhibit AKH release after a sugar-rich meal, to suppress further sugar feeding while promoting protein intake.
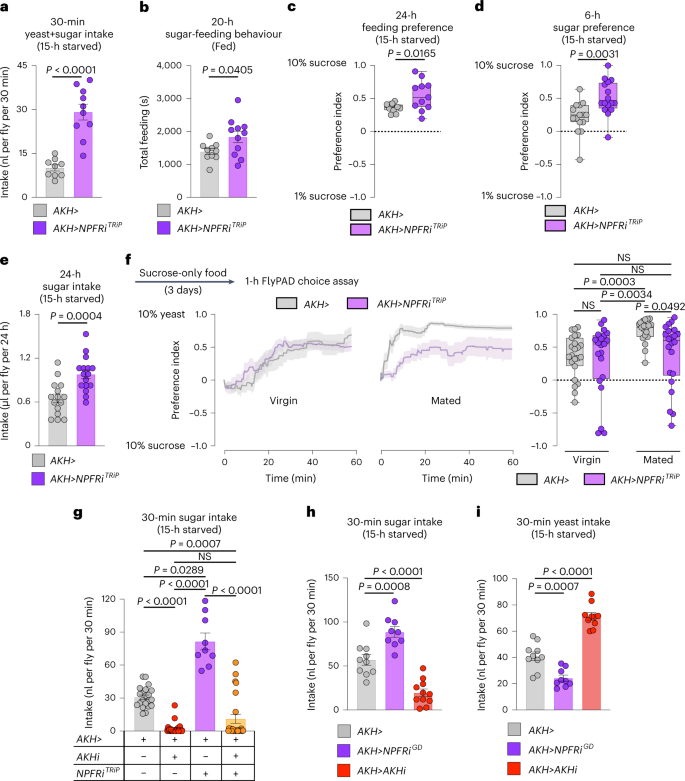
a, Food intake measured by dye assay; n = 9 AKH>, n = 10 AKH>NPFRiTRiP. b, Time spent feeding on sucrose measured by FLIC; n = 10 AKH>, n = 11 AKH>NPFRiTRiP. c, Behavioural preference for 1 versus 10% sugar solution measured by FLIC; both n = 11. d, Consumption preference for 1 versus 10% sucrose measured by CAFÉ assay; n = 15 AKH>, n = 16 AKH>NPFRiTRiP. e, Sucrose consumption measured by CAFÉ assay; n = 15 AKH>, n = 16 AKH>NPFRiTRiP. f, Behavioural preference using flyPAD. Lines represent means, and shading indicates s.e.m.; virgins n = 23 AKH>, n = 21 AKH>NPFRiTRiP; mated n = 19 AKH>, n = 22 AKH>NPFRiTRiP. g, Rescue of sugar overconsumption induced by NPFR knockdown in the APCs through simultaneous AKH knockdown, by dye assay; n = 21 AKH>, n = 24 AKH>AKHiKK, n = 9 AKH>NPFRiTRiP, n = 24 AKH>AKHiKK, NPFRiTRiP. h,i, Sucrose (h) and yeast (i) intake measured over 30 min by dye assay. h, n = 10 AKH>, n = 9 AKH>NPFRiGD, n = 11 AKH>AKHiKK. i, n = 10 AKH>, n = 9 AKH>NPFRiGD, n = 10 AKH>AKHiKK. All animals were mated females, except in f. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, median, 75th percentile and maximum values. NS, not significant. a–d, Two-tailed unpaired Student’s t-test. e, Two-tailed unpaired Mann–Whitney U-test. f,g, One-way Kruskal–Wallis ANOVA with Dunn’s multiple-comparisons test. h,i, One-way ANOVA with Dunnett’s multiple-comparisons test.
Source data
AKH regulates appetites for sugar and protein-rich food
AKH is described as a generic hunger hormone released during nutritional deprivation18,35,39. However, our findings indicate that AKH regulates food choice. To confirm this effect, we examined the feeding behaviour of mated AKH mutant females and found that these animals exhibited significantly reduced sugar intake (Fig. 8a), suggesting that AKH promotes sugar preference. On this basis, this we expected an increase in yeast preference with loss of AKH, so we used assay conditions in which animals do not normally exhibit strong yeast preference: 15-hour starvation rather than 3-day yeast deprivation. Loss of AKH, including adult-restricted APC-specific knockdown (with GAL80TS; AKH>, together referred to as AKHts>), led to a striking shift in preference towards yeast using the two-choice dye assay in mated females (Fig. 8b). Consistent with their increased intake of and preference for yeast food, mated AKH mutant females displayed a strong increase in the amount of time spent exploring patches of yeast food compared to sugar patches (Fig. 8c). This further supports a role for AKH in controlling feeding decisions, biasing behaviour towards sugar intake. Consistent with this, activation of the APCs to induce AKH release caused increased sugar intake while decreasing yeast intake, effects that were blocked by simultaneous AKH knockdown (Fig. 8d,e), indicating that they were mediated by AKH. Circulating sugar levels and whole-body TAG levels were not altered (Extended Data Fig. 6a,b), suggesting that the observed AKH-induced feeding phenotypes are direct effects that precede detectable metabolic changes. Together, these findings indicate that AKH is a hormone that controls selective feeding decisions by increasing appetite for sugar and reducing intake of protein food.
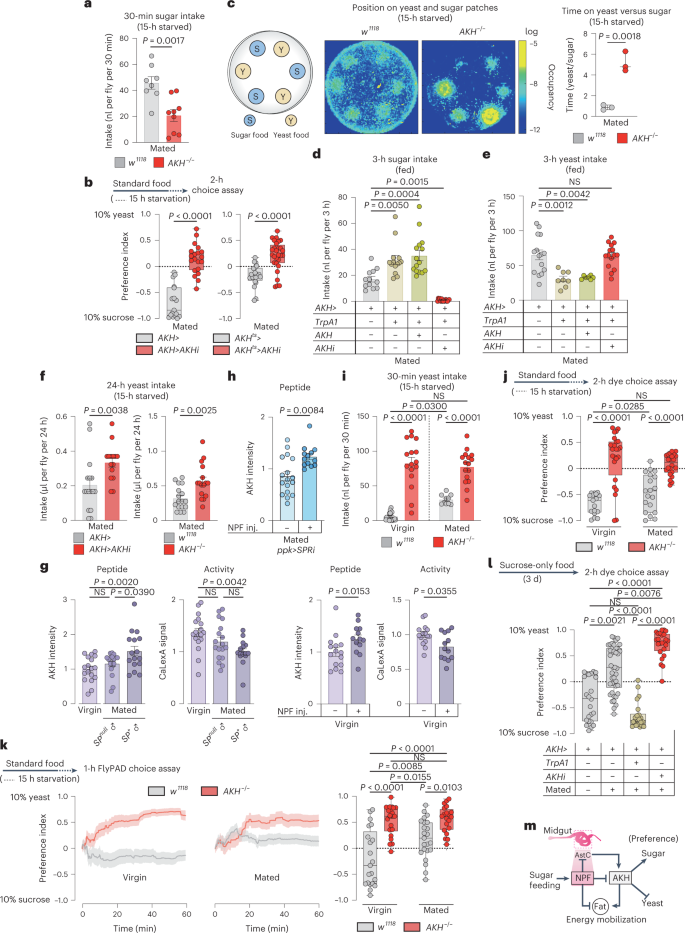
a, Sugar intake by dye assay. n = 8 w1118, n = 9 AKH−/−. b, Consumption preference, n = 17 AKH>, n = 20 AKH>AKHiKK, n = 20 AKHts>, n = 28 AKHts>AKHiKK. c, Heat map of 30 min tracking of 12 females per genotype. Ratio of time spent on yeast versus sugar patches, n = 3. d,e, Sugar and yeast intake using dye assay, at 29 °C for TrpA1 activation. d, n = 12 AKH>, n = 14 AKH>TrpA1, n = 15 AKH>TrpA1+AKH, n = 15 AKH>TrpA1+AKHi. e, n = 15 AKH>, n = 8 AKH>TrpA1, n = 7 AKH>TrpA1+AKH, n = 15 AKH>TrpA1+AKHi. f, Yeast intake measured by CAFÉ assay. n = 6 AKH>, n = 15 AKH>AKHiKK, n = 17 w1118, n = 16 AKH−/−. g, APCs staining and cell activity, left two panels show n = APCs from 18 w1118 virgins, n = 18 w1118 females mated to SP-deficient males (SP0/Df(3L)delta130), n = 16 w1118 females mated to SP+ males. Right two panels show n = 15 w1118 virgins without NPF injection, n = 13 w1118 virgins with NPF injection. h, AKH staining intensity in mated females with SPR knockdown in the ppk+ neurons with or without NPF injection, n = 17 ppk>SPRi mated females, n = 13 ppk>SPRi mated females with NPF injection. i, Yeast consumption measured by dye assay, n = 18 w1118 virgins, n = 16 AKH−/− virgins, n = 10 mated w1118 females, n = 16 mated AKH−/− females. j, Consumption preference using the two-choice dye assay; n = 17 w1118 virgins, n = 24 AKH−/− virgins, n = 19 mated w1118 females, n = 22 mated AKH−/− females. k, Cumulative behavioural preference using flyPAD. Lines represent means, and shading indicates s.e.m., n = 21 w1118 virgins, n = 22 AKH−/− virgins, n = 23 mated w1118 females, n = 20 mated AKH−/− females. l, Preference measured using the two-choice dye assay at 29 °C for TrpA1 activation, n = 23 AKH> virgins, n = 42 AKH>, n = 22 AKH>TrpA1, n = 24 AKH>AKHi mated females. m, A model of the NPF-AKH axis. Bars represent mean ± s.e.m. Box plots indicate minimum, 25th percentile, media, 75th percentile and maximum. Inj., injection; NS, not significant. a,b (right), c,f, Two-tailed unpaired Student’s t-test. b (left), g (right), h, Two-tailed unpaired Mann–Whitney U-test. d,e,g (left), i,k, One-way ANOVA with Tukey’s post hoc test. j, One-way ANOVA with Dunnett’s multiple comparison test or Kruskal–Wallis with Dunn’s multiple-comparisons test. l, One-way Kruskal–Wallis ANOVA with Dunn’s multiple-comparisons test.
Source data
AKH has recently emerged as a key factor in sex-specific metabolic regulation40. Unlike mated females, in which AKH loss enhanced yeast intake, males lacking AKH exhibited a decrease in yeast intake, indicating that AKH plays a sexually dimorphic role in feeding decisions (Fig. 8f and Extended Data Fig. 6c,d). Together these findings indicate that, in mated females, gut-derived NPF inhibits AKH secretion, and this inhibition suppresses sugar appetite and increases the consumption of protein-rich food. In this scenario, increased AKH signalling in virgins promotes preference for dietary sugar. We therefore examined whether mating reduces AKH release. Mating increased AKH peptide levels and reduced the calcium activity of the APCs, indicating repression of AKH release, and the effect on AKH was dependent on SP signalling (Fig. 8g). These findings suggest that SP signalling, through activation of gut NPF, contributes to the suppression of AKH release after mating. Injecting NPF peptide into virgin females increased AKH peptide levels within the APCs and reduced their calcium activity, indicating that NPF is sufficient to repress AKH secretion (Fig. 8g). Mating did not lead to AKH retention in females with knockdown of SPR in the Ppk+ neurons (Extended Data Fig. 6e), further indicating that SP-SPR signalling is required to repress AKH signalling after mating. To test whether NPF functions downstream of SP-SPR signalling in the AKH-regulatory hierarchy, we injected NPF into mated females with reduced SP-SPR signalling (ppk>SPR-RNAi) and found that this led to increased AKH peptide levels in the APCs (Fig. 8h). This suggests that NPF is sufficient downstream of the SP pathway to repress AKH signalling after mating, which increases yeast intake. We therefore conjectured that loss of AKH would increase yeast intake in virgin females, whereas increasing AKH signalling in virgin females would have little effect on feeding behaviour. To assess the effect of loss of AKH, we examined behaviour under starved conditions, under which AKH signalling is normally high. As expected, we found that while control females increased their yeast consumption in response to mating, similarly conditioned AKH mutant virgins displayed a striking overconsumption of yeast food that was not significantly altered by mating (Fig. 8i). Activation of the APCs to induce AKH release in the fed state, in which AKH signalling is generally lower, did not alter yeast intake in fed virgin females (Extended Data Fig. 6f), presumably because of the already higher AKH signalling in the virgin state, whereas as mentioned above this treatment did reduce the yeast intake of fed mated females (Fig. 8e). Consistent with these findings, virgin AKH mutant females exhibited a strong preference for yeast food that did not increase significantly in response to mating as it did in control females (Fig. 8j,k and Extended Data Fig. 6g). Together, this suggests that, in mated females, NPF acts through NPFR in the APCs to repress AKH signalling, which increases yeast intake. Consistent with this, we found that inducing AKH release was sufficient to block the high yeast preference exhibited by mated females after 3 days of yeast deprivation (Fig. 8l). To demonstrate that NPF regulates food choice via AKH, we injected NPF into mated females with APC-specific NPFR knockdown. This manipulation did not increase yeast preference after 3 days’ yeast deprivation (Extended Data Fig. 6h), suggesting that NPFR in the APCs is required for NPF to promote yeast preference. Furthermore, injection of NPF into AKH mutant virgin females also did not alter sugar versus yeast preference, indicating that AKH is required for mediating the effects of NPF-NPFR on food preference (Extended Data Fig. 6i). We propose that sugar-induced AKH-repressive NPF signalling from the gut constitutes a hormonal axis involved in suppressing sugar appetite and promoting intake of protein-rich food in mated females (Fig. 8m).